This week’s BEACON Researchers at Work blog post is by MSU postdoc Noah Ribeck.
 We are all taught the basic tenet of evolution by natural selection: occasionally an individual is born with a mutation that improves its chances of having more offspring. Anyone descended from this individual will then inherit this beneficial mutation, and so over generations, that mutation has a good chance of taking over the population.
We are all taught the basic tenet of evolution by natural selection: occasionally an individual is born with a mutation that improves its chances of having more offspring. Anyone descended from this individual will then inherit this beneficial mutation, and so over generations, that mutation has a good chance of taking over the population.
But when does evolution not quite work this way? Sometimes, the benefit of a mutation changes as it becomes more abundant in the population – this is called frequency-dependent selection. A classic example of this is Batesian mimicry, where “warning” systems of poisonous organisms (like distinctive coloration) are copied by non-poisonous species to gain the benefit of being left alone by predators. This strategy only works if relatively few are doing it: if there are too many mimics around, predators would learn not to be fooled by the tricky tactic.
In the lab, I study frequency-dependent selection in the Long-Term Evolution Experiment (LTEE), a set of 12 populations of E. coli that have been evolving in the lab of Richard Lenski for more than 58,000 generations. As you might imagine, there is nothing like Batesian mimicry here, but there is a relatively common mechanism of frequency dependence in microbes, called cross-feeding. This happens when one group feeds off a metabolic byproduct of another group. The cross-feeding group, then, only has an advantage when the group it’s feeding off is abundant. In other words, a mutation that confers the cross-feeding ability is advantageous only when rare. When this happens, the two groups can coexist in perfect harmony at an equilibrium ratio where each strategy works equally well.
The LTEE was started in 1988 with the purpose of being as simple as possible. There is only a single resource in the media (glucose) to prevent individuals from specializing on different energy sources. And to discourage cross-feeding, glucose is provided only at a low concentration to prevent a lot of buildup of metabolic byproducts. In other words, the LTEE was designed for E. coli to get better at using glucose, period.
And for the most part, that’s exactly what’s happened. But over the years, members of the lab have stumbled across a few different instances of long-term coexistence of multiple subpopulations – and in one case very long-term. One of the populations consists of two distinct subgroups that have been coexisting since around ~6,500 generations, just a few years into the experiment. They are affectionately called the “smalls” and “larges,” because the two groups were made obvious by the differently sized colonies they made on agar plates. We’ve also recently found an example of frequency dependence in another population, by looking at DNA sequence data of that population over time. In this case though, the coexistence of the two groups lasted only a few thousand generations.
To begin to study these examples, I’ve measured the competitive fitness (that is, a relative measure of reproduction speed) of the two groups against each other, at various different initial ratios. From this data, we expect to see that each group has an advantage (fitness>1) when rare, and a disadvantage (fitness<1) when common. And that’s exactly what happens:
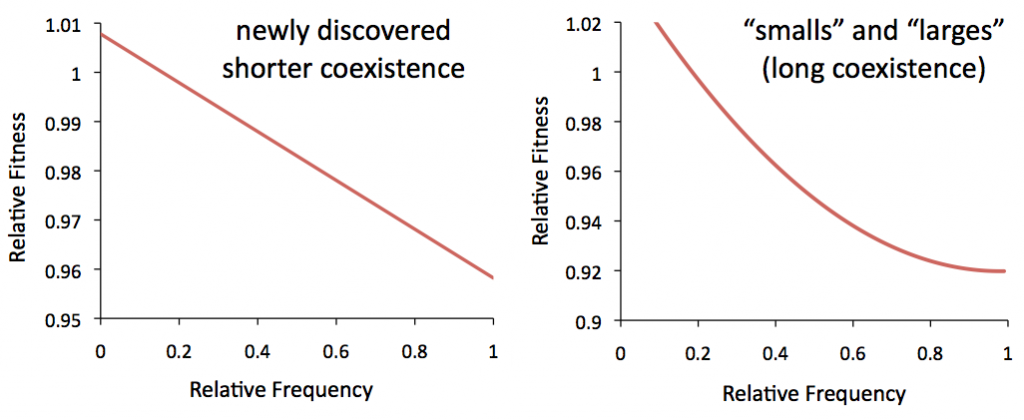
Measuring frequency-dependent fitness is a bit tricky, because we do it by measuring how fast one group grows while competing directly against the other. But when fitness is frequency-dependent, the fitness we’re trying to measure is continuously changing while we’re measuring it. We can account for this with the proper math, but that’s why the data you’re seeing here are smooth curves instead of data points.
To interpret this, we need to have some idea of what the frequency-dependence curve would look like for an actual cross-feeding interaction. To do that, we can model the ecology between the two groups mathematically, and compare that to our data. When we do that, the theoretical fitness a lot like a straight line, decreasing with frequency, very much like our recently discovered shorter coexistence:
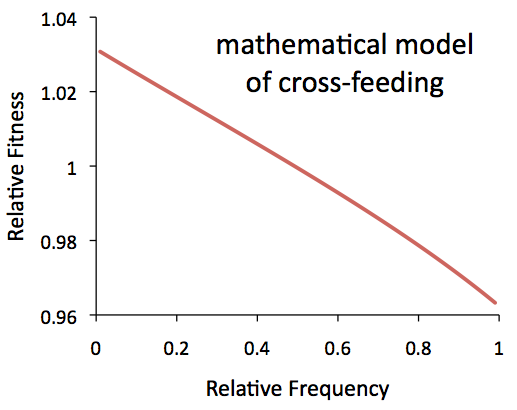
But the long-term coexistence (the smalls and larges) has a bit of a twist: their frequency-dependent fitness levels off a bit at when the smalls are common. So what’s going on with the smalls? Well, we actually already know that there’s something else happening there: when the smalls are common, they start to selectively kill off the larges! This means that the fitness of the smalls (relative to the larges) gets a boost when the smalls are abundant – in other words, the smalls have an added advantage when common. This flavor of frequency dependence is not sustainable on its own: if the benefit of a mutation gets bigger as it grows in frequency, it will simply drive the other group extinct. It’s only possible for this interaction to exist indefinitely because it’s paired with an offsetting disadvantage when common, most likely from cross-feeding.
I’m interested in the fundamental role of these ecological interactions in adaptation. Since frequency-dependent selection seems to be relatively common (even in an environment that was designed to avoid it!), can we rewrite some of the basic theories in population genetics to account for it? Do these ecologies serve to sustain genetic variance in populations to help them adapt faster? Do they serve as stepping stones to more highly beneficial genotypes that would have never been otherwise possible? Stay tuned as we work on these questions …
For more information about Noah’s work, you can contact him at ribeck at msu dot edu.